Research Article - Journal of Bacteriology and Infectious Diseases (2017) Journal of Bacteriology and Infectious Diseases (Special Issue 1-2017)
Analysis of antibody by Gamma function
Bin Zhao*, Aibing Li, Lichun Liang
College of Science, Northwest A&F University, Yangling, Shaanxi, China
- *Corresponding Author:
- Bin Zhao
College of Science
Northwest A&F University
Yangling
Shaanxi
China
Tel: +86 13028517572
E-mail: zhaobin835@nwsuaf.edu.cn
Accepted Date: October 30, 2017
Citation: Zhao B, Li A, Liang L. Analysis of antibody by Gamma function. J Bacteriol Infec Dis. 2017;1(1):1-8
Abstract
Along with the rapid development of genetic engineering technology and antibody engineering technology, humanized monoclonal antibody has been rapidly developed and gradually replaces the rat sourced monoclonal antibody. In this paper, we establish two new logarithmically completely monotonic functions involving the gamma function according to two preferred interaction geometries, necessary and sufficient conditions are presented for one of them to be logarithmically completely monotonic. As a consequence, a sharp inequality involving the gamma function is deduced to solve the problems of genetically engineered antibody.
Keywords
Gamma function, Genetically engineered antibody, Logarithmically completely monotonic, Inequality, Psi function
Introduction
Antibodies have been proven to be indispensable tools for biomedical applications. Different engineered antibodies have been development for various purposes according to the amino acid sequence and/or spatial structure of protein (Figure 1). At present, it is still difficult to predict the optimal structure of antibodies. Topology knowledge can be important in antibody application as well as transformation. Theoretically, we can obtain desired antibodies by using protein/gene engineering technology. For instance, we can transform the complementarity determining region (CDR) to promote the affinity of the antibody to antigen. Similarly, we could also transform any domain of antibody to make it bind with any desired target. Under this vision, topology is a powerful tool to predict the structure of protein and it will serve to antibody engineering. Our present work tries to explain, and predict, if possible, the change of structure, size and function of antibodies as well as their fragments from a topological perspective.

Figure 1: Different antibody formats. a: different antibody or engineered antibodies; b: different shape of antibody.
For Re (z) > 0 the classical Euler’s gamma function G and psi (digamma) function y are defined by
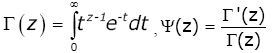 (1.1)
(1.1)
respectively. The derivatives ψ(n)(z) for n ∈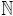 are known as
polygamma functions.
are known as
polygamma functions.
For ψ(z) [1], the following series representations are established:
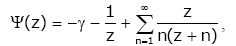 (1.2)
(1.2)
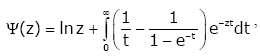 (1.3)
(1.3)
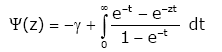 (1.4)
(1.4)
Where, g = 0.577215664901...denotes the Euler’s constant.
We next recall that a function f is said to be completely monotonic on an interval I , if f has derivatives of all orders on I which alternate successively in sign, that is,
(−1)nf(n) (x) ≥ 0 , (1.5)
for all x ∈ Iand for all n ≥ 0 . If inequality (1.5) is strict for all x ∈ Iand all n ≥ 0 , then f is said to be strictly completely monotonic [2-5]. The classical Bernstein–Widder theorem [6] states that a function f is completely monotonic on (0, ¥) if and only if it is a Laplace transform of some nonnegative measure m , that is,
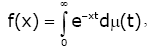 (1.6)
(1.6)
Where, m (t ) is non-decreasing and the integral converges for x > 0.
We recall also that a positive function f is said to be logarithmically completely monotonic on an interval I if f has derivatives of all orders on I and
(−1)n [ln f(x)](n) ≥ 0, (1.7)
for all x ∈ I and for all n ≥ 1. If inequality (1.7) is strict for all x ∈ I and all n ≥ 1, then f is said to be strictly logarithmically completely monotonic [7-9].
The antibody structure will be changed when it binds certain target (Figure 2a), i.e.,: antigen, receptor. How to describe the changes in the view of topology? The following cases will explain it in detail. It was proved explicitly by Berg C [8] and other articles that a logarithmically completely monotonic function must be completely monotonic.

Figure 2: Model of pH-dependent conformational change of FcRY and structures for the FcRY monomer and dimer. a: FcRY has an extended conformation at pH 8 (s*= 7.2 S) with no predicted interaction between the CysR-FNII domains and the CTLDs. At pH 6 the CysR-FNII region folds back and binds to the CTLDs, resulting in a more compact conformation (s*= 7.9 S) that is able to bind IgY. b: Likely orientations of FcRY and FcRY–IgY on a membrane. The two FcRY monomers on the right are shown in an orientation that would allow formation of a 2:1 FcRY–IgY complex.
Anderson GD et al. [10] proved that the function
g( x) = x(lnx − ψ( x)) (1.8)
is strictly decreasing and strictly convex on (0, ¥), with two limits
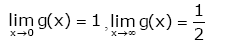 (1.9)
(1.9)
From (1.9) and the monotonicity of g ( x), then the double inequalities
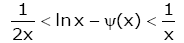 (1.10)
(1.10)
holds for all x > 0 .
By using the well-known Binet’s formula, Alzer H [11] generalized the monotonicity and convexity of g (x), that is, the function
ga(x) = xa(lnx − ψ(x)) (1.11)
is strictly completely monotonic on (0, ∞) if and only if α ≤ 1
Kershaw D and Laforgia A [12], proved that the function [Γ (1 + 1 / x)]x is decreasing on (0, ∞) and x[Γ (1 + 1 / x)]x is increasing on (0, ∞). These are equivalent to the function [Γ (1 + 1 / x)]1/x being increasing and [Γ (1 + 1 / x)]1/x / x being decreasing on (0, ∞) , respectively.
F Qi and Chp Chen [13] generalized these functions. They obtained the fact that for all x > 0 the function xr [Γ (1 + x)]1/x is strictly increasing for r ≥ 0 and strictly decreasing for r ≤ −1, respectively.
After the papain digestion, the remained antibody functional part (usually the Fab domain), will be smaller and the structure is also changed (Figure 1b). These changes can be revealed vividly using topology. Recently Qi F et al. [14] established another excellent result, which states that for given y ∈(−1, ∞) and α ∈(−∞, ∞) , let
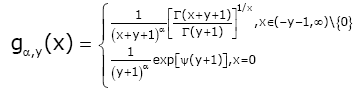 (1.12)
(1.12)
The function (1.12) is logarithmically completely monotonic with respect to x ∈(−y − 1, ∞) if and only if α ≥ max{1,1 / ( y + 1)} and if α ≤ min{1,12 / ( y + 1)}, the reciprocal of the function (1.12) is logarithmically completely monotonic with respect to x ∈(−y − 1, ∞) .
Antibodies occur spontaneously gathering and forming dimer, polymer, which will influence their functions (Figure 2b). In antibody engineering practice, it urgently needs some measures to overcome this difficulty. From topology perspective, we could understand this issue as follow.
Stimulated by the above results, we put forward the function as follows: for given y ∈(0, ∞) and real number a , let the function fα,y (x) be defined by:
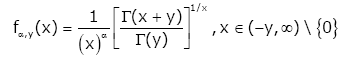 (1.13)
(1.13)
Our first result is contained in the following theorem.
Theorem 1. For the function (1.13), then the following statements are true:
(1) For any given y≥1, the function (1.13) is strictly logarithmically completely monotonic with respect to x Î(- y, ∞) \ {0}if and only if α ≥1 ;
(2) For any given 0 < y < 1, if α ≥ e−(1− y) / y , then the function (1.13) is strictly logarithmically completely monotonic with respect to x ∈(− y,∞) \ {0};
(3) For any given y > 0, the reciprocal of the function (1.13) is strictly logarithmically completely monotonic with respect to x ∈(− y,∞) \ {0} if and only if α ≤ 0.
Our second result is presented in the following theorem.
Theorem 2. For any given y ∈[1,∞] , let the function hy ( x) be defined on (0, ∞) by
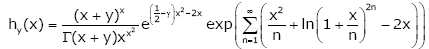 (1.14)
(1.14)
Where γ denotes the Euler’s constant, then the function (1.14) is strictly logarithmically completely monotonic with respect to x on (0, ∞).
The following corollary can be derived from Theorems 2 immediately.
Corollary 1. For any given y ≥ 1, the inequality
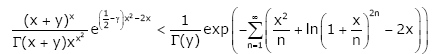 (1.15)
(1.15)
holds for all x > 0.
Lemma
In order to prove our main results, we need the following lemmas.
It is well known that Bernoulli polynomials Bk (x)and Euler polynomials Ek (x) are defined by
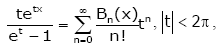
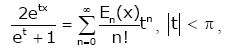 (2.2)
(2.2)
respectively. The Bernoulli numbers Bn are denoted by Bn = Bn (0) , while the Euler numbers En are defined by En = 2 En (1/2).
The following summation formula is given:
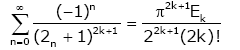 (2.3)
(2.3)
for any nonnegative integer k , which implies
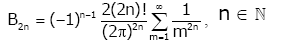 (2.4)
(2.4)
In particular, it is known that for all n Î we have
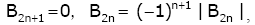 (2.5)
(2.5)
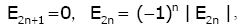 (2.6)
(2.6)
And the first few nonzero values are
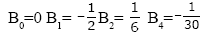
E0 = 1, E2 = -1, E4 = 5,
The Bernoulli and Euler numbers and polynomials are generalized [15-21].
Lemma 1. For real number x > 0 and natural number m [22,23], then
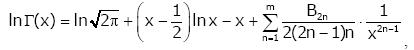
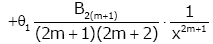
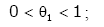 (2.7)
(2.7)
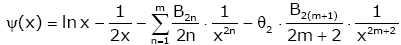 ,
, 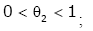 (2.8)
(2.8)
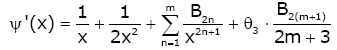 ,
, 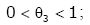 (2.9)
(2.9)
 (2.10)
(2.10)
Remark 1. θ1 , θ2 , θ3 , θ4 only depend on natural number m.
Lemma 2. For real number x > 0 and natural number n, we have [24]:
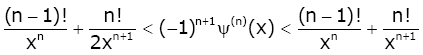 (2.11)
(2.11)
Lemma 3 For real number x > 0 and natural number n, we have [1,17]:
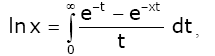 (2.12)
(2.12)
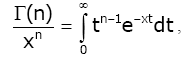 (2.13)
(2.13)
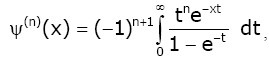 (2.14)
(2.14)
Lemma 4. Let the sequence of functions un (x) for n ∈ be
defined on (0, ∞) by
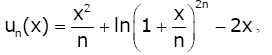 (2.15)
(2.15)
the series 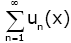 is differentiable on (0, ∞), that is,
is differentiable on (0, ∞), that is,
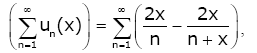 (2.16)
(2.16)
Proof. It is obvious that 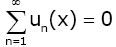 , therefore
, therefore 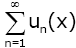 converges at x = 0. In order to prove (2.16), we need only to
show that the inner closed uniform convergence of the series
converges at x = 0. In order to prove (2.16), we need only to
show that the inner closed uniform convergence of the series 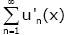 on (0, ∞). From (2.15), we have
on (0, ∞). From (2.15), we have
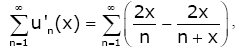 (2.17)
(2.17)
For any interval [a,b] ⊂ (0,∞) , we have
 (2.18)
(2.18)
x ∈[a,b] It is easy to check that the series 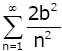 converges, which and Weierstrass M-test implies that the series
converges, which and Weierstrass M-test implies that the series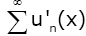 is inner closed uniformly convergent on (0, ∞).
Hence the series
is inner closed uniformly convergent on (0, ∞).
Hence the series 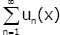 is differentiable on (0, ∞) and the
identity (2.17) holds for x ≥ 0 .
is differentiable on (0, ∞) and the
identity (2.17) holds for x ≥ 0 .
The lemma is proved.
Lemma 5. For 0 < α ≤1 and real number b, let the function Qa,b ( x) be defined by
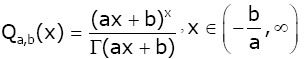 (2.19)
(2.19)
If b ≥ a, then the function (2.19) satisfies
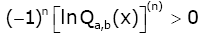 (2.20)
(2.20)
for all 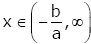 and n = 2,3,….
and n = 2,3,….
Proof. Taking the logarithm of Qa,b ( x) yields
lnQ (x) = xln(ax + b) −l nΓ(ax + b), (2.21)
and differentiating lnQa,b (x) , then
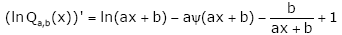 (2.22)
(2.22)
For given integer n ≥ 2, we get
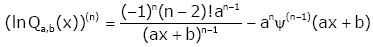
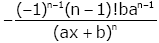 (2.23)
(2.23)
and, by the identities (2.13) and (2.14), (2.23) can be written as
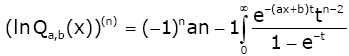 (2.24)
(2.24)
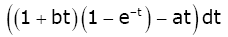
Let
ρ(t) = (1 + bt)(1 − e−t ) −at and q(t) = (1 + t)(1 − e−t ) −t .
It is easy to check that
q' (t ) = te−t > 0, t ∈(0, ∞) (2.25)
therefore q (t ) is strictly increasing on (0, ∞), and then q (t ) > q (0) = 0.
The following two cases will complete the proof of Lemma 5.
Case 1. If 0 < a ≤ 1 ≤ b, then since q (t ) > 0 for t > 0, we have at a t < (1+ t )(1- e-t ) £ (1+ bt )(1- e-t ), (2.26)
which implies 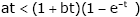 , and then p (t ) > 0
for all t > 0.
, and then p (t ) > 0
for all t > 0.
From (2.24), we know that the inequality (2.20) holds for x ∈(−b a,∞) and integer n ≥ 2.
Case 2. If 0 < a ≤ b ≤ 1, then we get
p' (t ) = b − a + e−t (bt + 1 − b) ≥ bte−t > 0, t ∈(0, ∞) (2.27)
Therefore p (t ) is strictly increasing on (0, ∞), and then p (t ) > p (0) = 0 .
From (2.24), we know that the inequality (2.20) holds for x ∈(-b a , ∞) and integer n ≥ 2.
The lemma is proved.
Proof of Theorems
Proof of Theorem 1. For x ≠ 0and natural number n, taking the logarithmically differential into consideration yields
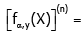
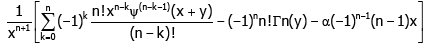 (3.1)
(3.1)
where ψ(−1) (x + y) and ψ(0) (x + y) stand for lnΓ(x + y) and ψ(x + y) respectively.
Furthermore, differentiating 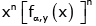 directly gives
directly gives
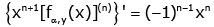
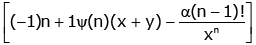 (3.2)
(3.2)
Making use of (2.11) and (2.13) shows that for all n ∈ and any fixed y > 0 ,the double inequality
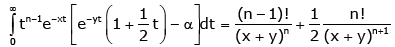
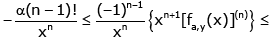
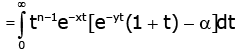 (3.3)
(3.3)
holds for all x ∈(− y,∞) \ {0} and α∈ (−∞,∞) .
For any fixed y ∈(0,∞), let u (t ) and v (t ) be defined on (−∞,∞) by
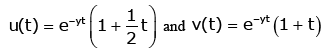 respectively.
respectively.
Differentiating u (t ) and v (t ) directly, we obtain
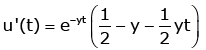 (3.4)
(3.4)
v'(t) = e−yt (1 − y − yt) (3.5)
Therefore, for given y ∈(0,∞)we have
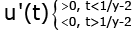 (3.6)
(3.6)
and
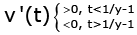 (3.7)
(3.7)
From (3.6) and (3.7), we conclude that for all t > 0 we obtain u(t) > 0, (3.8)
and
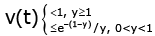 (3.9)
(3.9)
From (3.3) and (3.8)-(3.9), it is easy to see that
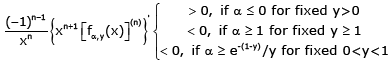 (3.10)
(3.10)
For all n ∈ and all x∈ (−y,∞) \ {0}.
On the one hand, if x ∈(0, ∞), then the inequalities (3.10) can be equivalently changed into
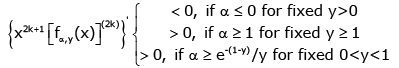 (3.11)
(3.11)
and
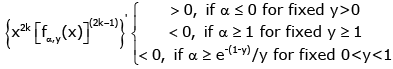 (3.12)
(3.12)
for k ∈ .
From (3.1), then simple computation shows that 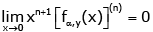 (3.13)
(3.13)
for all n ∈ and any given y ∈(0,∞). As a result,
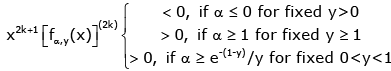 (3.14)
(3.14)
and
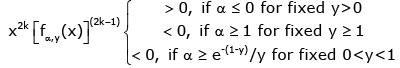 (3.15)
(3.15)
for all k ∈ and all x > 0.
Therefore, (3.14) and (3.15) imply
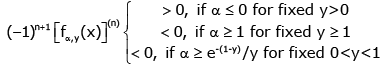 (3.16)
(3.16)
for all n ∈ and all x > 0.
Hence, if either α ≥ e−(1− y) / y for given 0 < y < 1 or α ≥ 1 for given y ≥ 1, the function (1.13) is strictly logarithmically completely monotonic with respect to x on (0, ∞), and if α ≤ 0 for given y > 0, so is the reciprocal of the function (1.13).
On the other hand, if x ∈(− y, 0) for any given y > 0, then (3.10) implies
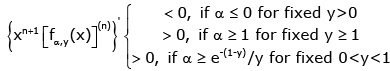 (3.17)
(3.17)
for n ∈.
In view of (3.13), we can conclude that
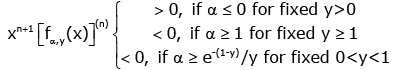 (3.18)
(3.18)
for n ∈ . It is obvious that (3.18) is equivalent to that (3.14)
and (3.15) hold for any given y > 0 and x ∈(- y,0).Therefore, it
is easy to prove similarly that (3.16) is also valid on x ∈(- y,0)
for any given y > 0 and all n ∈ .
The amino acid of antibody/protein possesses different preferences. Thus we can conduct site-directed mutation to promote the affinity and/or hydrophilic with the prediction of topology. For example, bovine antibodies have an unusual structure comprising a β-strand ‘stalk’ domain and a disulphidebonded ‘knob’ domain in CDR3 (Figure 3). Attempts have been made to utilize such amino acid preference for antibody drug development.

Figure 3: Unique structural domain in bovine IgG antibodies and application
Consequently, the function (1.13) is the same logarithmically completely monotonicity on (- y,0) as on (0, ∞), that is, if either α ≥ e–(1−y) / y for given 0 < y < 1 or α ≥ 1for given y ≥ 1 the function (1.13) is strictly logarithmically completely monotonic with respect to x on (- y, 0) , and if α ≤ 0 for given y > 0, so is the reciprocal of the function (1.13).
Conversely, we assume that the reciprocal of the function (1.13) is strictly logarithmically completely monotonic on (− y,∞) \ {0} for any given y > 0. Then we have for any given y > 0 and all x > 0
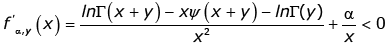 (3.19)
(3.19)
which implies
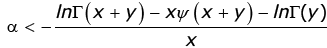 (3.20)
(3.20)
By L’Hˆospital’s rule, we have 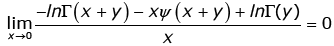 (3.21)
(3.21)
for any given y > 0. By virtue of (3.20) and (3.21), we conclude that the necessary condition for the reciprocal of the function (1.13) to be strictly logarithmically completely monotonic is α ≤ 0.
If the function (1.13) is logarithmically completely monotonic on (− y,∞) \ {0}
for any given y > 0, then the inequality (3.19) and (3.20) are reversed for any given y > 0 and all x > 0.
By utilizing (2.7) and (2.8), it is easy to see that
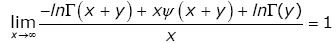 (3.22)
(3.22)
for any given y > 0. In fact, it is not difficult to show that the necessary condition for the function (1.13) to be strictly logarithmically completely monotonic is α ≥ 1.
The proof of Theorem 1 is completed.
Proof of Theorem 2. Taking the logarithm of hy ( x) gives
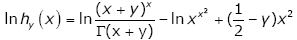
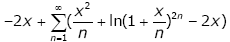 (3.23)
(3.23)
Let
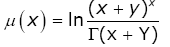 (3.24)
(3.24)
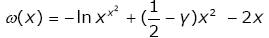
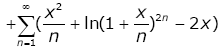 (3.25)
(3.25)
then
lnhy (x) = μ(x) + ω(x) . (3.26)
In view of Lemma 4, straightforward calculation gives
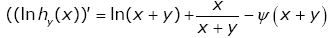
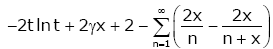
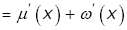 (3.27)
(3.27)
By virtue of (1.2), the identity (3.27) is equivalent to
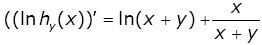
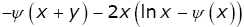
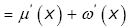 (3.28)
(3.28)
By Lemma 5, we know that μ'(x) is strictly increasing on (0, ¥), which and (1.10) imply the limit of μ'(x) equals 1 as x → ∞ therefore
μ'(x) < 1 (3.29)
holds for all x > 0.
We know that g ( x) is strictly completely monotonic on (0, ¥), where g ( x) defined by (1.8), hence for given integer n ≥ 0 , the inequality.
(−1)n+1 (ω' ( x))(n) > 0 (3.30)
holds for all x > 0.
And then by using inequality (1.9) and (1.10), we get
−2 < ω' ( x) < −1 (3.31)
for all x > 0 .
From (3.29) and (3.31), we conclude that
(lnhy( x)) = μ( x) +ω( x) < 0 (3.32)
for all that x > 0. Utilizing Lemma 5 and (3.30), for given integer n ≥ 2, it is easy to see that equation for all x > 0.
Theorem 2 follows from (3.32) and (3.33).
Thus the proof of Theorem 2 is completed [25-31].
Conclusion
In conclusion we establish two new logarithmically completely monotonic functions involving the gamma function according to two preferred interaction geometries, and a sharp inequality involving the gamma function is deduced to solve the problems of genetically engineering antibody. It is necessary to address, many other aspects (such as thermal condition, alkalinity or acidity, adhesion of antibody) are also playing key roles in antibody functioning, which could be also understood from bio-mathematical perspective, and such knowledge will be in return useful for biomedical application of antibodies as well as proteins [25-31].
Acknowledgements
This work was supported by the Ph.D. programs foundation of ministry of education (20130204110023); the national nature science foundation (31572556); the international scientific and technological cooperation and exchange program in Shaanxi province (S2015YFKW0002), and the Key Construction Program of International Cooperation Base in S&T, Shaanxi Province (2015SD0018), China.
References
- Erdélyi A, Magnus W, Oberhettinger F, et al. Higher Transcendental Functions. New York: McGraw-Hill. 1953.
- Bernstein S. Sur les fonctions absolument monotones. Acta Math. 1929;52:1–66.
- Hausdorff F. Summationsmethoden und Momentfolgen I. Math. 1921;9:74–109.
- Qi F, Chen P. A complete monotonicity property of the gamma function. J Math Anal Appl. 2004;296(2):603–607.
- Widder DV. Necessary and sufficient conditions for the representation of a function as a La- place integral. Trans Amer Math Soc. 1931;33:851–892.
- Widder DV. The Laplace Transform. Princeton University Press, Princeton. 1941.
- Atanassov RD, Tsoukrovski UV. Some properties of a class of logarithmically completely monotonic functions. C R Acad Bulgare Sci. 1988;41:21–23.
- Berg C. Integral representation of some functions related to the gamma function. Mediterr J Math. 2004;1:433–439.
- Grinshpan AZ, Ismail MEH. Completely monotonic functions involving the gamma and q-gamma functions. Proc Amer Math Soc. 2006;134:1153–1160.
- Anderson GD, Barnard RW, Richards KC, et al. Inequalties for zero-balanced hypergeometric functions. Trans Amer Math Soc. 1995;347:1713–1723.
- Alzer H. On some inequalities for the gamma and psi functions. Math Comp. 1997;66:373–389.
- Kershaw D, Laforgia A. Monotonicity results for the gamma function. Atti Accad Sci To- rino Cl Sci Fis Mat Natur. 1985;119:127–133.
- Qi F, Chen Chp. Monotonicity and convexity results for functions involving the gamma function. J Appl Math Sci. 2004;1:27–36.
- Qi F, Wei CF, Guo BN. Complete monotonicity of a function involving the ratio of gam- ma functions and applications. Banach J Math Anal. 2012;6:35–44.
- Guo BN, Qi F. A property of logarithmically absolutely monotonic functions and the loga- rithmically complete monotonicity of a power-exponential function. Politehn Univ Bucharest Sci Ser A Appl Math Phys. 2010;72(2): 21–30.
- Wang Z-X, Guo DR. Introduction to special function. The Series of Advanced Physics of Peking University. Beijing, China: Peking University Press. 2000.
- Abramowitz M, Stegun IA. Handbook of Mathematical Functions with Formulas, Graphs, and Mathematical Tables. Washington: Applied Mathematics Series, National Bureau of Standards. 1970;55(9).
- Guo BN, Qi F. Generalization of Bernoulli polynomials. Int J Math Educ Sci Technol. 2002;33:428–431.
- Luo QM, Guo BN, Qi F, et al. Generalizations of Bernoulli numbers and polynomials. Int J Math Math Sci. 2003:3769–3776.
- Luo QM, Qi F. Relationships between generalized Bernoulli numbers and polynomials and generalized Euler numbers and polynomials. Advanced Studies in Contemporary Mathematics (Kyungshang). 2003;7:11–18.
- Luo QM, Qi F, Debnath L. Generalizations of Euler numbers and polynomials. Int J Math Math Sci . 2003:3893–3901.
- Qi F, Guo BN. Monotonicity and convexity of ratio between gamma functions to different powers. Journal of the Indonesian Mathematical Society. 2005;11:39–49.
- Fichtenholz GM. Differential- und Integralrechnung. Berlin: VEB Deutscher Verlagder Wissenschaften. 1964;2.
- Guo BN, Qi F, Two new proofs of the complete monotonicity of a function involving the psi function. Bull Korean Math Soc. 2010;47:103–111.
- Scott AM, Wolchok JD, Lloyd J. Antibody therapy of cancer. Nature Rev. 2012;12:278- 287.
- Carlos F, Lucia R, Jeffrey W. High-affinity self-reactive human antibodies by design and selection: Targeting the integrin ligand binding site. Medical Sciences. 1993,90:10003-10007.
- Miroslava O, Xavier E. Human genetics moves from clinic to bench - and back. Genome Biology. 2005;3:343.
- Repp R, Hvan Ojik H, Valerius T, et al. Phase I clinical trial of the bispecific antibody MDX-H210(anti-FcgRI anti-HER-2/neu) in combination with Filgrastim(G-CSF) for treatment of advanced breast cancer. Br J Cancer. 2003,89:2234-2243.
- Alinari L, Lapalombella R, Andritsos L, et al. Alemtuzumab (Campath-1H) in the treatment of chronic lymphocyticleukemia. Nature. 2007;26:3644- 3553.
- Burns JC, Mason WH, Hauger SB, et al. Infliximab treatment for refractory Kawasaki syndrome. J Pediatr. 2005;146(5):662-667.
- Baloch AR, Baloch AW, Sutton BJ, et al. Antibody mimetics: Promising complementary agents to animal-sourced antibodies. Crit Rev Biotechnol. 2014;36(2):268-275.