- Journal of RNA and Genomics (2010) Research Report
RNAi effects on actin mRNAs in Homalodisca vitripennis cells
| Radkin Honzak*,Vera Strunzova,Janouskova L,Moravcova M Psychiatric Hospital Bohnice, Prague 8, 18100 Ústavní 91, Czech Republic |
| Corresponding Author: Radkin Honzak M.D, Ph.D, Psychiatric Hospital Bohnice, Prague 8, 18100 Ústavní 91, Czech Republic E-mail: radkinh@seznam.cz Tel: +420-224917479 |
| Received: 23 September 2014 Accepted: 11 February 2015 Published: 21 March 2015 |
Abstract
The xylem feeding leafhopper Homalodisaca vitripennis (H. vitripennis) is an unusually robust and efficient vector of Xylella fastidiosa, a Gram-negative bacterium which causes several very important plant diseases. Here we investigated RNA interference (RNAi) to target actin, a key component of insect cells and whole bodies, in H. vitripennis cells. RNAi effectors were delivered via lipid based transfection and real-time RTPCR, RNA hybridization, and microscopic analyses were employed to verify RNAi effects. When actin dsRNAs were used, a 10-fold decrease in the target H. vitripennis actin mRNA level was seen in cells. Altered phenotypic effects also were evident in transfected cells, as were small interfering RNAs, hallmarks of RNAi. The use of H. vitripennis cells and RNAi offers new opportunities to research hemipterans, the most important insect vectors of plant pathogens.
Keywords |
| Homalodisca vitripennis, RNA interference, glassy-winged sharpshooter, Pierce’s disease |
Introduction |
| The glassy-winged sharpshooter (GWSS), Homalodisca vitripennis (Germar) (H. vitripennis, formerly H. coagulata) is among the most robust and threatening vectors of the xylem limited Gram-negative bacterium Xylella fastidiosa (X. fastidiosa) (Hopkins and Purcell, 2002). Strains of this bacterium cause Pierce’s disease, a devastating disease occurring in winegrapes from California to Texas to Florida, phony peach disease in the southern United States, oleander and almond leaf scorch in California and Texas, and citrus variegated chlorosis in Brazil (Redak et al, 2003). Since these diseases are invading new areas, new approaches to control H. vitripennis and other insect vectors are needed. |
| RNA interference (RNAi) has been suggested as strategy to develop “insect-proof plants” (Gordon and Waterhouse, 2007; Huvenne and Smagghe, 2009), but practical applications of this technology are yet to be developed. However, fundamental RNAi-based studies are proving to be very valuable (Mao et al, 2007; Baum et al, 2007). Intrathoracic injection of dsRNAs is the most effective way to induce RNAi in whole insects (Price and Gatehouse, 2008), but it is laborious, relatively slow and not applicable to all insects. Therefore, insect cell line systems might prove to be beneficial for RNAi studies, offering a means for rapid, efficient, fundamental and even practical studies. For example, cell line-based screens have been used to identify RNAi effects in human (Yang et al, 2002), African green monkey (Cercopithecus aethiops) (Pacca et al, 2009) and the fruit fly (Drosophila melanogaster) (Cheng et al, 2005) (and http://flyrnai.org) cell lines. |
| We used the H. vitripennis -Z15 cell line (Kamita et al, 2005), and evaluated specific H. vitripennis RNAs as potential RNAi inducers. Our effort demonstrates for the first time that RNAi activity is not only present in a leafhopper species, but is inducible in H. vitripennis cells. Furthermore, we were able to induce RNAi effects against actin, a major component of all eukaryotic cells (Doherty and McMahon, 2008). Our results suggest that RNAi can be used as fundamental tool to better understand H. vitripennis genetics and physiology, and possibly for investigating the dynamics of ‘plant-pathogen-vector’ interactions. |
Materials and Methods |
| H. vitripennis -Z15 cell culture and transfection |
| The H. vitripennis -Z15 cell line was maintained as described by Kamita and colleagues (Kamita et al, 2005). Five x 105 cells per 35mm diameter culture dish in logarithmic growth phase were transfected with 2μg of dsRNA (Cellfectin Transfection Reagent, Invitrogen), and were harvested 24, 48 and 72hrs post transfection. Transfection reagent was used as control. |
| DsRNA production |
| H. vitripennis actin (region 1-1000 on sequence AY588061), sar1 (region 3-556 on sequence AY588076), arginine kinase (region 1-1000 on sequence AY588062), Lian-Aa1 (HM104624) and synthetic GFP (Lindbo, 2007) cDNAs, were cloned in both orientations downstream the T7 promoter in pGEM-T Easy (Promega). The plasmids served as templates for dsRNA transcription (MEGAscript, Ambion). |
| Real time RT-PCR validation. |
| Total RNA was extracted from transfected H. vitripennis- Z15 cells, 72 post transfection (RNeasy Mini Kit, Qiagen). Real time PCR primers as given below were designed using sequences AY588061 AY588076, and AY588062, respectively:- |
| actinf 994: 5’GCTCCCCCTGAGAGGAAGTA |
| actinr 1066: 5’TCTGTTGGAAGGTGGACAGA |
| sar1f 100: 5’ACCACCTTGCTTCACATGCT |
| sar1r 176: 5’TTCTGAGGTGGGGTGTAAGG |
| argkinhf 980: 5’CTGAAGGCGGAGTGTATGACATC |
| argkinhr 1039: 5’ACAGCCTGGTACTCGGTGAGA |
| Real time RT-PCR reactions were performed as previously described (Pignatta et al, 2007). Data analyses were performed by comparative CT or ΔΔCT method (Applied Biosystems). |
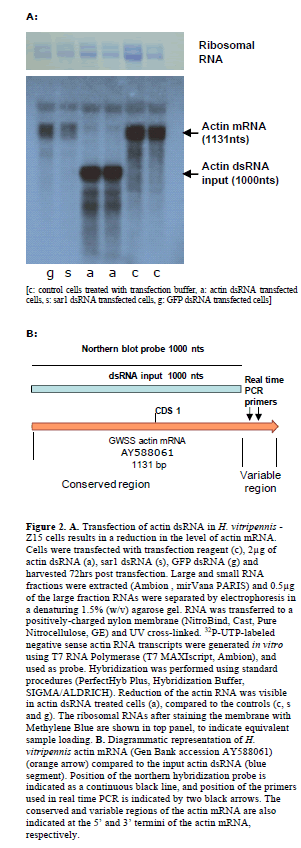
| Northern blot analysis of H. vitripennis -Z15 cells |
| Cells were transfected with transfection reagent, 2μg of actin, sar1, or gfp dsRNAs as above and harvested 72hr post transfection. Large and small RNA fractions were extracted (mirVana PARIS, Ambion) and separately analyzed by northern hybridization (Jacob-Wilk et al, 2009). 32PUTP-labeled negative sense actin RNA transcripts (region 1-1000 on sequence AY588061) were generated in vitro using T7 RNA Polymerase (T7 MAXIscript, Ambion), and used as a probe. Hybridization was performed using standard procedures (PerfectHyb Plus, Hybridization Buffer, SIGMA/ALDRICH, and mirVana PARIS, Ambion). |
| Actin visualization |
| H. vitripennis cells were grown to logarithmic phase in 35 mm-diameter culture dishes containing a glass coverslip, and transfected as above. Coverslips were removed 72hr post transfection. Actin staining of the transfected cells was performed using Alexa Fluor 488 phalloidin, (Molecular Probes, Invitrogen) and cells were observed by fluorescence microscopy (Leica DM5000B). |
Results |
| Only a few H. vitripennis cDNA sequences corresponding to putative proteins are currently available in GenBank (DQ118408, AY588060, to AY588069, AY588071, AY58807 4 to AY588076, AY869766), thus limiting the targets for RNAi studies. Of these sequences a putative muscle actin (AY588061) and a putative cytoplasmic actin (AY588060) were available. These two sequences share 85% overall nucleotide identity, and regions of complete identity are found at the mRNA 5’-termini thereby facilitating the design of RNAi experiments that target both sequences. Because actin is one of the major components of the cell cytoskeleton and of muscle cells, we believed it could be a good potential target to investigate H. vitripennis cell biology. We also investigated RNAi effects on H. vitripennis sar1 mRNA (AY588076). Sar1 orthologs are known to bind to the ER and are connected to actin filaments, where they are involved in transport from the endoplasmic reticulum to the Golgi (Kuge et al, 1994), suggesting their mRNAs would be suitable as references in real time RT-PCR analyses. Real time RT-PCR was adopted first as the tool to measure the relative amounts of corresponding mRNAs in H. vitripennis -Z15 cells. Because no ribosomal RNA or beta tubulin sequences (these are those most commonly used as reference genes for real time PCR assays) were available in Genbank for H. vitripennis, the sar1 and actin were alternately used as both reference and target genes. |
| Potential RNAi effectors were introduced into H. vitripennis -Z15 cells via lipid-based transfection. Viability tests showed no obvious differences among transfected cells, and mortality during the experimental time frame (72 hrs) was extremely low, on the order of 1 to 2% (data not shown), demonstrating absence of toxicity of the transfection reagent and RNAi effectors to H. vitripennis -Z15 cells. In initial experiments, we observed that following transfection, target mRNA accumulation decreased over time from 24 to 72hrs (data not shown). In addition, among the different types of RNAi inducers tested by us, dsRNA was the most effective (data not shown), and we found that a minimal dose of dsRNA (equal to 2μg of dsRNA in 5x105 cells per 35mm-diameter culture dish) was needed to efficiently induce RNAi effects. Based on these results, we used dsRNA as the RNAi inducer in the experiments described here, and we tested RNAi effects at 72hr time point. |
| H. vitripennis -Z15 cells were transfected with dsRNAs specific for H. vitripennis arginine kinase (AY588062) (Wang et al, 2009), actin, sar1 or Lian-Aa1 (a non-LTR retrotransposon) as well as for the green fluorescent protein (GFP), or with transfection reagent alone. At 72hr time point, realtime RT-PCR analyses showed that actin mRNA was decreased by 95% in cells transfected with actin dsRNA, in comparison to mock-transfected control cells (Figure 1). However, the other dsRNAs also induced some reduction (~50-60%) in actin mRNA relative to the mock control. When these same cells were analyzed for the sar1 mRNA, the sar1 dsRNA transfected cells had 75% reduction relative to the transfection reagent control, but in contrast to results for actin mRNA cells transfected with the other dsRNAs showed approximately twice the amount of sar1 mRNA relative to the control cells. Thus off-target effects were seen for both actin and sar1 analyses. If control transfection reagent treated cells are not considered and only dsRNA treated cells are compared, then reductions of 87% for the actin mRNA and 87% for the sar1 mRNA were seen when compared to the control GFP dsRNA transfected cells. |
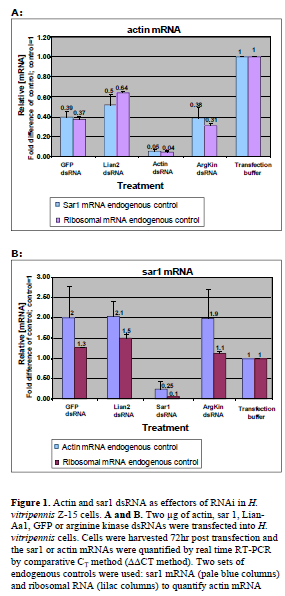 |
| (panel A); actin mRNA (blue columns) and ribosomal RNA (red columns) to quantify sar1 mRNA (panel B). RNAi effects are shown by the sar1 and actin mRNA reductions in sar1 and actin dsRNA transfected cells respectively, compared to cells treated with transfection reagent only (calibrators). The percentage knockdown for actin mRNA was 95% relative to the transfection reagent control, and 87% compared to the GFP control. For sar1 percentage knockdown was 75% relative to the transfection reagent and 87% compared to the GFP control (left columns in panels A and B respectively), when we used sar1 and actin mRNA as endogenous controls. Similar results were obtained using ribosomal RNA as endogenous controls. Numbers above the columns represent the fold differences of mRNA concentration compared to the control. Experiments were repeated three times. Of these, the cell transfection was performed twice in three replicates and once in two replicates. Real time RT-PCR samples were always loaded in duplicates. Error bars above the columns indicate the standard deviation among the 3 experiments. |
| In order to determine if the off-target effects seen in our experiments were the result of a real time RT-PCR artifact, we used two additional real time RT-PCR endogenous controls, the H. vitripennis putative arginine kinase and ribosomal RNA. The partial H. vitripennis ribosomal RNA sequence was obtained by us analyzing H. vitripennis EST sequences deposited in GenBank. In these experiments, reductions of the targeted (actin and sar1) mRNAs and offtarget effects similar to those described above were found (Figure 1), confirming the validity of our RT-PCR methodology. |
| We next used northern hybridization analysis to assess levels of mRNAs. An obvious decrease in the levels of the actin mRNA was seen in H. vitripennis cells transfected with actin dsRNA in comparison to H. vitripennis -Z15 cells that were transfected with reagent only or sar1 or GFP dsRNAs (Figure 2), thus supporting the RT-PCR data. The probe used for northern hybridization was designed for the conserved region of the actin mRNAs, and exactly to match the dsRNA used as RNAi inducer. Because of the homology among the 5’ regions of the cytoplasmic and muscle actin mRNAs, we expected to silence both actin sequences, resulting in a reduction of both mRNAs. In contrast, the primers used for real time RT-PCR analysis were designed on the variable region of the muscle actin mRNAs. This choice was necessary in order to avoid detecting the input transfected actin dsRNA in real time RT-PCR analysis. The results obtained by northern hybridization and by real time RT-PCR both demonstrate the specific reduction of actin mRNA in the actin dsRNA transfected cells, but because these analyses were designed to detect different regions in the actin mRNAs, the results cannot be quantitatively compared. Similar to the RT-PCR analysis, in the northern hybridization cells transfected with dsRNA GFP (g) and sar1 (s) showed a decrease in actin mRNA when compared to cells transfected with transfection reagent only (c) (Figure 2). |
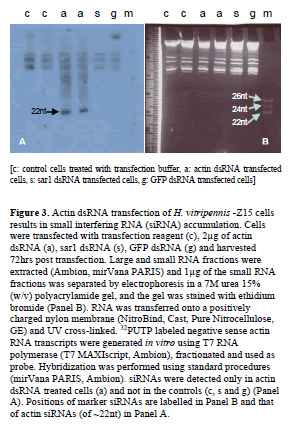
| RNA interference is not only manifested by a decrease in the target mRNA accumulation, but also by the appearance of small interfering RNAs (siRNAs) corresponding to mRNA target sequence (Tomoyasu et al, 2008). Therefore the presence of siRNAs specific for actin mRNA was investigated by northern hybridization analysis. Actinspecific siRNAs were seen only in RNAs extracted from actin dsRNA transfected cells (a) and not in the controls (c, s and g) (Figure 3). Comparison of the positions of the actin siRNAs with those of oligonucleotide markers shows that the actin siRNAs were approximately 21nt. Taken together these data clearly showed that RNAi was induced in the H. vitripennis -Z15 cells. |
 |
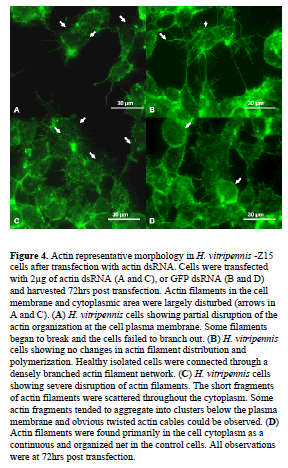
| Because actin is an abundant and essential component of eukaryotic cells, and actin filaments can be specifically labeled with fluorescent phallotoxin derivatives, such as phalloidin, we determined whether actin dsRNA treated cells showed phenotypic alterations that were consistent with the downregulation of actin mRNA accumulation. Microscopic observation of H. vitripennis -Z15 cells transfected with actin dsRNAs showed clear alterations in cell morphology during the experiment time frame (72 hrs) in comparison to control transfected cells (Figure 4). In actin dsRNA transfected cells, actin filaments were not apparent at the cell membranes at 72hr time point, especially at the cell periphery. The actin filaments were distorted and appeared broken in the cytoplasm, and cells were not connected to each other. Actin filaments often failed to develop and appeared to be shorter (Figure 4, panels A and C). In control cells transfected with GFP dsRNA (Figure 4, panels B and D), dense actin filaments were observed on the plasma membrane at the cell periphery, and tight junction strands formed welldeveloped networks. |
 |
 |
Discussion |
| Here we show that specific RNAi effects can be induced in the H. vitripennis -Z15 cell line. Transfection of dsRNA molecules directed towards mRNAs of endogenous genes reduced mRNA levels dramatically, as was shown by real time RT-PCR analysis and northern hybridization. To confirm that the mRNA reductions seen were due to RNAi, we also clearly showed the presence of RNAi hallmarks, gene-specific siRNAs, and thus confirmed achieving RNAi in H. vitripennis cells. In addition to the actin-specific siRNAs induced here (see Figure 3), we also found that when GFP-specific dsRNAs were introduced into GWSS Z15 cells, GFP-specific siRNAs were generated (data not shown), demonstrating that processing of dsRNA occurs even in the absence of a mRNA target for small RNA amplification. H. vitripennis sequences of genes and proteins involved in the RNAi pathways are still lacking from the public databases, but another member of the Hemiptera, the pea aphid Acyrthosiphon pisum, posses proteins similar to Dicer 1, required for siRNA-directed mRNA cleavage in Drososphila (Lee et al, 2004) and for Argonaute 2b, required for successful siRNA response in Drososphila (Obbard et al, 2006) suggesting that other organisms belonging to the order of the Hemiptera (such as H. vitripennis) are likely to have RNAi homologs. |
| We also were able to achieve actin-specific phenotypic RNAi effects on cultured H. vitripennis cells. Actin isoforms are found in all eukaryotic organisms and both actin genes and proteins are highly conserved in different species, where the expression of actin is time and tissue regulated. Actin is a crucial component for cell motility, muscle contraction, cell division and growth processes, cell signalling, cell shape and junctions, and it is an ideal candidate for fundamental RNAi studies. We only saw cell mortality of 1-2% in our experiments and this level is very similar to that seen for the transfection control. However, because we used only the 72hr time point for our analysis we cannot conclude whether or not cell mortality might be higher if cells were analyzed at later times. Furthermore, H. vitripennis -Z15 population doubling time is less than 72hrs, rendering it difficult to estimate the mortality due to dsRNA transfection. |
| Although we observed significant reductions in target mRNAs, we also saw off-target effects when we compared dsRNA-treated cells to those treated only with the transfection reagent control (mock treatment). At this time we do not know the reason for these observations. We used long dsRNA molecules as RNAi inducers, and it might be possible that short regions of the dsRNAs could be homologous to other mRNA sequences in H. vitripennis. We used long dsRNAs instead of short siRNAs because dsRNA induced greater mRNA reduction than other forms of RNAi inducers in our preliminary experiments (data not shown). Others also have shown that dsRNAs seems to be more effective for inducing RNAi effects in insects (Huvenne and Smagghe, 2009). Furthermore, even using single siRNAs does not guarantee the absence of off-target effects. Extensive off-target effects in silencing experiments are well known in mammalian systems (Jackson et al, 2003), and are likely to be observed in insect systems as well. The mechanism underlying these off-target effects is not yet clear, since the introduction of siRNAs (even at low concentrations) into mammalian cells causes not only the down regulation but also the up regulation of unrelated genes (Tschuch et al, 2008). The biological role of RNAi is hypothesized to originate as a cellular response to RNAs of viral origin (Van Rij, 2008) resulting in the triggering of cytoplamic RNA silencing (Baulcombe, 2004). The above mentioned cellular responses could be speculated to be part of a defense mechanism (Hammond et al, 2000; Adelman et al, 2002). It would be of interest to verify if RNAimediated cell defense increases trafficking between the Golgi and the ER (sar1 is involved in this pathway) and reduces cell division (actin is a cell structural component) leading to increased accumulation of sar1 mRNA and decreased actin mRNA. |
| In addition to fundamental studies, our hope is that RNAi for insects such as H. vitripennis might have potential practical application. Xylella fastidiosa is an important bacterial pathogen of economically important crops like grape, citrus, and almond (Redak et al, 2003). RNAi-based efforts directed toward the control of insect plant pests are now becoming potentially feasible (Gordon and Waterhouse, 2007). Since actin is also a key component in whole insects, where it is expressed as muscle actin and as cytoskeletal actin, it might represent a reasonable RNAi target in whole insects. Nevertheless, H. vitripennis-Z15 cells offer a new tool for RNAi studies toward hemipterans, the most important vectors of plant pathogens. |
Conclusions |
| We show here for the first time that RNAi effects can be induced in H. vitripennis cells. From this we conclude that these cells offer an immediate and valuable genetic system for fundamental studies on H. vitripennis and other leafhoppers, and can be used as a model to elucidate RNAi or other metabolic pathways, such as those involving actin, specific to leafhoppers. Considering the importance of leafhoppers and other hemipterans to agriculture, we believe that H. vitripennis -Z15 cells also have potential for practical studies. |
Acknowledgements |
| This research was supported by a grant from the UC Pierce’s Disease Research Grant Program. We would like to thank MR Sudarshana for helping to start this project, HC Tsui for precious and technically challenging lab assistance, D Starr for feedback on actin properties, Bryony Bonning for donating the H. vitripennis -Z15 cell line, L Stewart for reviewing this manuscript and useful comments, and B Hammock for allowing the use of the tissue culture facilities. |
Compteting Interests |
| None declared |
List of Abbreviations |
| H. vitripennis: Homalodisca vitripennis |
| GWSS: Glassy-winged sharpshooter |
| PD: Pierce’s disease |
References
|