- Journal of RNA and Genomics (2010) Research Report
Evaluation of RNA-knockdown strategies for modulation of cytochrome P450 reductase activity in mouse hepatocytes
| Neil CW Mackenzie1, Simon G Lillico1, Ken Brown2, Charles R Wolf2,3 and Christopher BA Whitelaw1* 1Division of Developmental Biology, The Roslin Institute and Royal (Dick) School of Veterinary Studies, University of Edinburgh, Roslin, Midlothian, EH25 9PS, UK 2CXR Biosciences Limited, James Lindsay Place, Dundee Technopole, Dundee, DD1 5JJ, UK 3University of Dundee Biomedical Research Institute, Ninewells Hospital and Medical School, Dundee, DD1 9SY, UK |
| Corresponding Author: Neil Mackenzie, Email: neil.mackenzie@roslin.ed.ac.uk, Tel: +44 (0)131 5274200, Fax: +44 (0)131 4400434 |
| Received 05 August 2010; Revised 02 November 2010; Accepted 18 November 2010; Published online 26 November 2010 |
| © Copyright The Authors: This is an open access article, published under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/). This license permits noncommercial use, distribution and reproduction of the article, provided the original work is appropriately acknowledged with correct citation details. |
Abstract
Transgenic technologies can provide important animal models for studying drug-metabolizing enzymes. Our overall aim was to generate versatile cell and animal systems that exhibited varying levels of cytochrome P450 oxidoreductase (POR) activity, more accurately modelling the human population for pharmacological and toxicology studies. Towards this goal we evaluated RNA-interference constructs designed for use in vitro and in vivo for reducing POR activity in hepatocytes. This study clearly demonstrates that both POR protein level and reductase activity can be significantly knocked down in Hepa-1 cells in vitro, while highlighting the difficulty in predicting knockdown efficiency in transgenic animals. The high levels of embryonic lethality observed, and inability to produce multi-copy transgenic animals indicates that high levels of shRNA expression may be detrimental to embryonic development.
KEYWORDS |
| RNA-interference, drug metabolism, lentivirus vectors, liver metabolism, transgenic mice |
INTRODUCTION |
| The metabolism of drugs leading to functional modification or degradation occurs primarily in the liver. This process represents an important determinant of the duration and intensity of the pharmacological action of drugs (Thummel et al, 1997), with outcomes ranging from complete detoxification to the production of metabolites that are more toxic than the original drug. Phase I reactions can either activate or inactivate a drug through a variety of biochemical reactions, with the majority of oxidative events occurring through the cytochrome P450 (CYP) family of enzymes (Nebert et al, 1996). Phase II reactions often function on Phase I generated metabolites, and involve drug-conjugating enzymes that usually result in detoxification (Sheenan et al, 2001). |
| The liver represents a major site of Phase I drug metabolism through the function of the CYP enzymes (Nebert et al, 1996). The significant variability in drug clearance and clinical response observed between individuals often reflects differences in CYP activity (Ingelman-Sundberg, 2004). The CYP monoxygenases represent a large family of genes that are broadly classified into two groups (Nelson, 1999); those involved in drug metabolism and xenobiotic clearance, which are present at variable levels in different individuals, and those required for specific biological pathways including cholesterol biosynthesis, retinoic acid metabolism and steroid biosynthesis. All CYP enzymes are activated by NADPH:Cytochrome P450 Oxidoreductase (POR; E.C.1.6.2.4). POR catalyses the transfer of electrons from NADPH to CYP enzymes and in the absence of POR, CYP enzymes are essentially inactive. The interaction of drugs with CYP enzymes, and subsequent alteration of CYP activity, is a major source of adverse drug reactions. |
| Transgenic technologies can provide important animal models for studying drug-metabolizing enzymes, specifically, animals can be engineered to enable studies on the regulation of enzymatic expression and the impact of variable levels of enzyme activity on pharmacological action and associated toxicity. The multiple genes that encode CYP enzymes, and their overlapping functions, make the analysis of gene knockout studies difficult (Henderson et al, 2003). An alternative approach has been to knockout POR, thereby affecting all CYP activity. The POR-null mutation in mice is embryonic lethal (Shen et al, 2002; Wu et al, 2005), but conditional knockout strategies have shown the value of this approach to understanding drug metabolism (Henderson et al, 2003; Finn et al, 2007). Mice lacking hepatic POR are viable and fertile, but completely devoid of liver CYP activity, and are unable to metabolize testosterone, acetaminophen and pentobarbital (Henderson et al 2003). With the aim to develop transgenic strategies providing intermediate POR activities, in this study we have evaluated RNA-interference (RNAi) approaches to knockdown POR activity in the mouse liver. |
MATERIALS AND METHODS |
| Vector construction |
| shRNA vectors were obtained from Sigma expressing various short hairpin sequences under the control of the human ubiquitous U6 pol-III promoter. |
| Target sequences: |
| shPOR1: 5'CCTGACCTACTGGTTCATCTT |
| shPOR2: 5'CGGAGGCACATCCTAGCCATT |
| shPOR3: 5'GCATCTAATGCACCTGGAATT |
| shPOR4: 5'CGGGAAGGAACATTATTGTAT |
| shPOR5: 5'GCTCGAAATATGGCCAAAGAT |
| These vectors contained a puromycin resistance gene under the control of a PGK promoter. This was removed using Nsi I and Bam HI, the backbone was blunted and a blunt ended eGFP fragment was ligated into the plasmid using T4 DNA ligase. |
| Lentivirus production |
| Self-inactivating (SIN) lentivirus was generated by Fugene-6 (Roche) mediated co-transfection of the three plasmids encoding the required packaging proteins, envelope and viral genome – 12μg psPAX2, 6μg VSV-G and 9μg pLKO-shRNA, repectively – a T150 flask seeded the previous day at 1x107 cells per flask (Al Yacoub et al, 2007). Conditioned medium was removed at 24hr and 48hr, filtered and centrifuged at 7,000xg O/N at 4°C, the resultant pellet was resuspended in 5ml TSSM buffer consisting of 20mM Tris, pH 7.3, 100mM sodium chloride, 10mg/ml sucrose and 10mg/ml mannitol, and concentrated by ultra-centrifuge at 20,000xg for 2hr at 4°C. The viral pellet was then resuspended in 100μl TSSM and aliquoted for storage at -80°C. Virus titres were determined by Polybrene-mediated serial dilution transduction of HT1080 cells and GFP expressing positive colonies were counted after 5 days (Al Yacoub et al, 2007). Concentrated viral titres ranged from 3.8x109 TU/ml to 6x109 TU/ml. |
| Cell culture and lentivirus transduction |
| Hepa-1 cells were grown at 37°C (5%, v/v, CO2) in Dubelecos modified Eagle medium (DMEM) supplemented with 10% (v/v) foetal calf serum (FCS, Gibco). Cells were grown to 80-90% confluence before being passaged. Hepa-1 cells were seeded at ~12000 cells per well in a 24-well plate 24hr prior to transduction with lentivirus at a multiplicity of infection (MOI) of ~1 in the presence of 8μg/ml polybrene. The cells were then FACS sorted to isolate the cells into which the pro-virus had integrated and maintained as a pooled population for no more than 10 passages. |
| POR expression and functional assays |
| Microsomal protein extracts were purified from Hepa-1 cells and mouse tissue using Sigma Endoplasmic Reticulum Extraction kit following the manufacturer’s protocol. Protein concentration was determined using BCA protein assay kit (Thermo Scientific) and expression was analysed by western blot employing ECL detection. POR detection was through rabbit CH59 anti-POR polyclonal antibody (kind gift from Colin Henderson, Dundee University: Henderson et al, 2003) and horseradish peroxidise labelled polyclonal goat anti-rabbit antibody (Dako). POR levels were normalised to β-actin using anti-mouse β-actin (Sigma) and goat anti-mouse (Dako) antibodies. The intensity of the bands was then quantified using PhosphorImager and QuantityOne software. The reductase activity of microsomal POR was determined using a colorimetric Cytochrome P450 reductase assay kit (Sigma) following the manufacturer’s protocol (Plonne et al, 1999). |
| Generation of transgenic mice |
| All animal experimentation was performed after review by Animal Ethics Committee and under UK Home Office Licence. Transgenic mice were generated as described previously (Ritchie et al, 2007). Eggs harvested from super-ovulated C57BL6 X CBA females were injected with a 10x or 1x pulse of ~50pl of 109 TU/ml1080 virus using standard micro-injection procedure. The virus was injected into the peri-vitellin space using a glass microinjection needle. Following injection the eggs were transferred to M16 medium (Sigma) for incubation at 37°C 5% (v/v) CO2 until day 4.5 where 15-18 developed blastocysts were surgically implanted into a psuedopregnant female. The resultant pups were genotyped by Southern blot (Whitelaw et al, 1992) and PCR (Vasey et al, 2008), using either: |
| eGFP-F: 5’CACATGAAGCAGCACGACTT with |
| eGFP-R: 5’TCTGGGTGGACAGGTAGTGG; or |
| HIVpsiF: 5’GAGAGAGATGGGTGCGAGAAG with |
| HIVpsiR: 5’GCTGTGCGGTGGTCTTACTT |
| as previously described. Transgenic mice were bred with C57BL6 X CBA F1 stock mice. |
RESULTS |
| Evaluation of shRNA vectors in vitro |
| POR knockdown was analysed by western blot in shPOR1 (72%), shPOR4 (55%) and shPOR5 (49%) transduced cell populations although given the variance between replicates only the reduction observed in shPOR1 cells was statistically significant (Figure 1). To determine if the shRNA knockdown of POR protein levels influenced hepatic cellular function we performed a reductase assay (Plonne et al, 1999). The lowest reductase levels were consistently observed for shPOR1 and shPOR5, reflecting the POR expression levels both showing a significant reduction in P450 reductase activity (Figure 1). The relatively high variance observed between replicates could be a consequence of varying numbers of viral integration events between the examined samples. A subsequent experiment, where cells were sorted according to GFP expression, indicated that those cells expressing higher levels of GFP grew slower than those expressing lower levels of GFP and when re-analysed by FACS their GFP expression profile had changed (data not shown). Higher GFP expression was assumed to be as a result of a greater number of integration events, and thus it may be that these cells also express higher quantities of shRNA. This suggests that there may be a significant reduction in growth rate or even toxicity associated with high numbers of integration events and transgene expression, indeed toxicity has previously been reported in cells expressing high levels of shRNAs (Fish et al, 2004). This shift in population may have affected different experimental replicates to a greater or larger degree resulting in a high variance in expression and activity read-outs. |
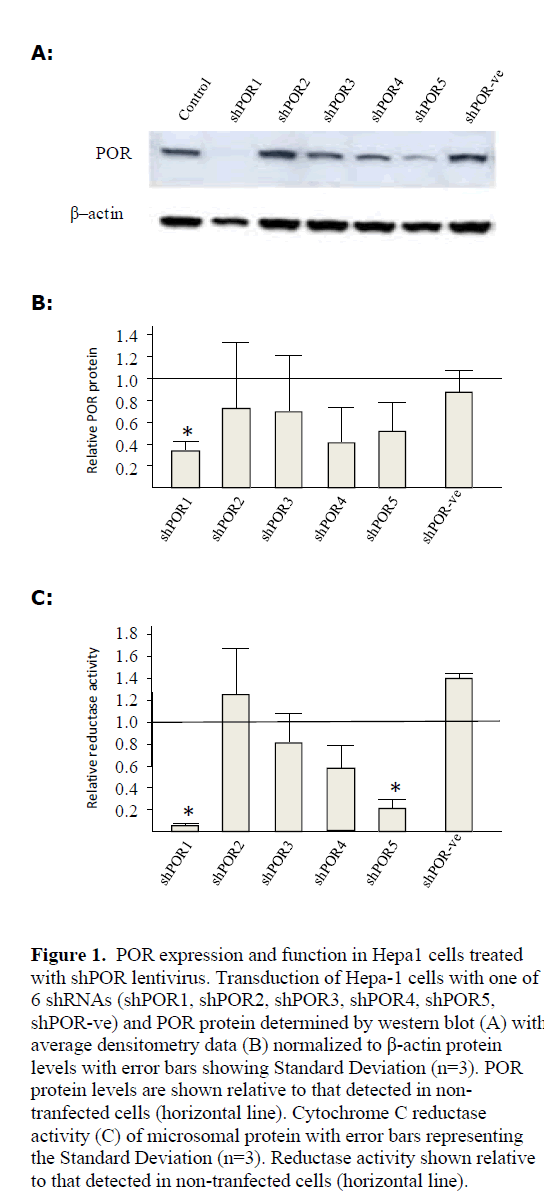 |
| Evaluation of shRNA vectors in vivo |
| shRNAs shown to give different levels of knockdown were used to generate transgenic animals. We attempted two separate peri-vitelline injection regimes in an attempt to get various levels of POR knockdown depending on transgene copy number; 1x50pl and 10x50pl of high-titre lentivirus (at least 3.8x109 TU/ml), the outcome of all the injections carried out are summarised in Table 1. Initial development of embryos to blastocysts showed a considerably lower level of development in the 10x injection batch (1x50pl: 241/376 (64%); 10x50pl: 143/375 (38%)) in all viral injections. This indicates that high concentrations of viral particles and/or shRNA expression reduce blastocyst development irrespective of target sequence. Integrational mutagenesis is a recognised problem of using lentiviruses however studies using similar injection strategies showed no significant reduction in development where up to 21 transgene integration events were seen (Lois et al, 2002; Ritchie et al, 2007), indicating that this may be a symptom of high levels of shRNA expression. |
| 1x50pl injection of shPOR1 into developing zygotes yielded three founder transgenic mice that were identified as positive by PCR of tail clip biopsy DNA, Southern blot analysis showed each positive contained only one copy of the shPOR1 transgene (data not shown); offspring were bred from two of the founder animals and genotyped by Southern blot (Figure 2A) and PCR. Liver microsomal protein extracts from the off-spring were analysed by western blot and Cytochrome P450 reductase assay (Figure 3). A clear relationship between POR expression and reductase activity was observed in these mice (Figure 3C) but, overall, no significant difference in POR expression or reductase activity was observed when compared to wild-type (WT) littermates. |
| Curiously all transgenic mice successfully raised carried only one copy of the transgene (Figure 2); similar studies have shown both higher efficiency of transgenesis and the generation of multicopy transgenic animals using similar quantities of virus (Lois et al, 2002). It is conceivable that in our study, any multicopy animals, which would presumably have higher shRNA expression levels and (based on our in vitro data) reduced POR activity, may have died in utero. Reduced growth rate seen in Hepa-1 cells, reduced blastocyst development and the following observations in the animal studies support this scenario. |
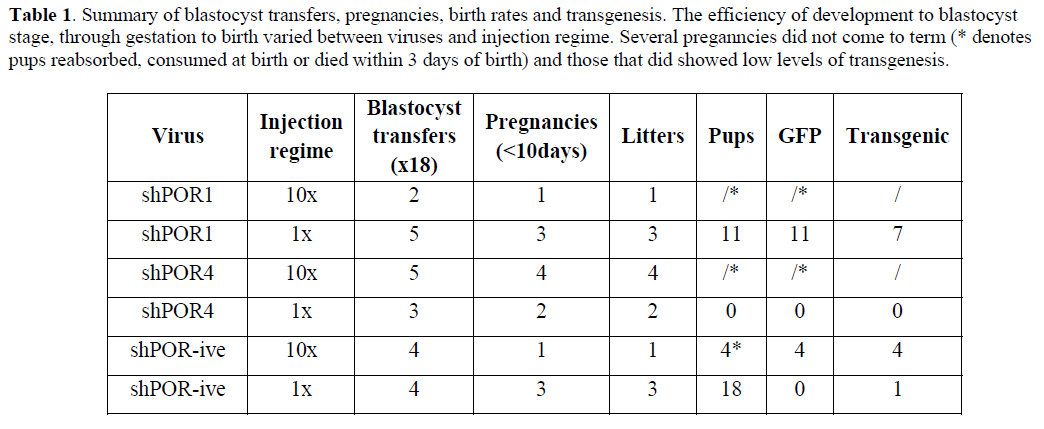 |
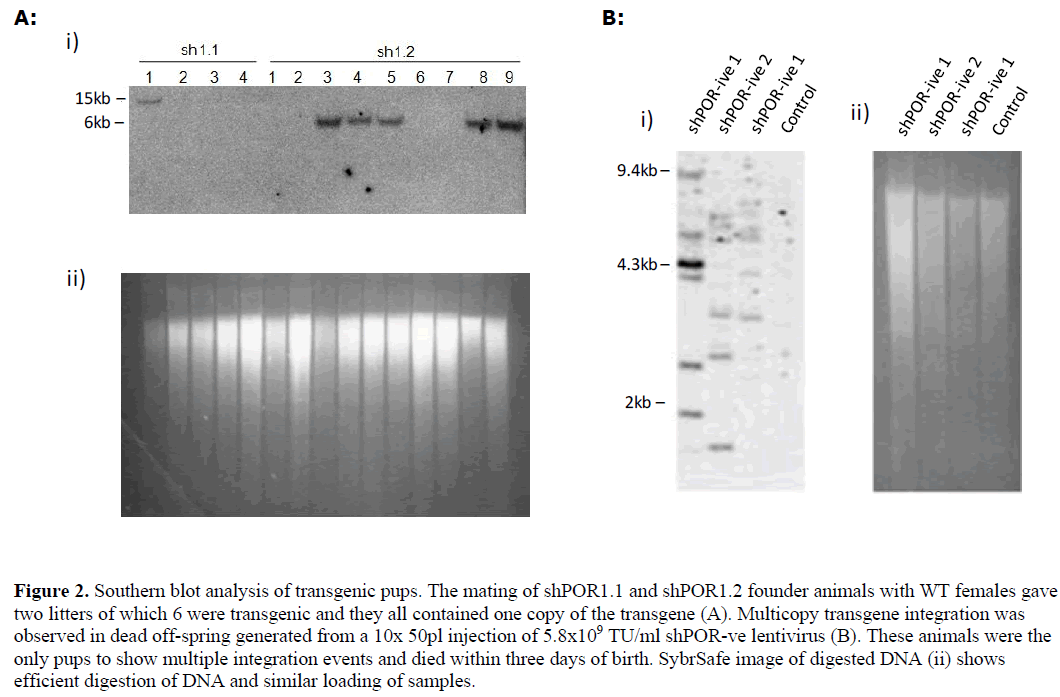 |
| In all 10x injections the pups were either re-absorbed or consumed at birth by the mother, in the one litter that survived (shPOR-ve) all the pups had died within 3 days. Where DNA samples were isolated from dead animals, Southern blot data indicated multiple copies of the transgene (Figure 2). The majority of 1x injections led to successful pregnancies although a low level of transgenesis was obtained (8/29; 27.5%) and all transgenic animals only contained one copy of the transgene (Figure 2b). For shPOR lentivirus injections less than half identified pregnancies (at day 10 gestation) resulted in births. If embryonic lethality was occurring it could result from high levels of shRNA (irrespective of target) which has been reported to cause mortality due to competitive inhibition of the endogenous miRNA system (Castanotto et al, 2007; Gimm et al, 2007) or reflect the known phenotypic consequence of significantly reducing POR activity during development (Shen et al 2005). |
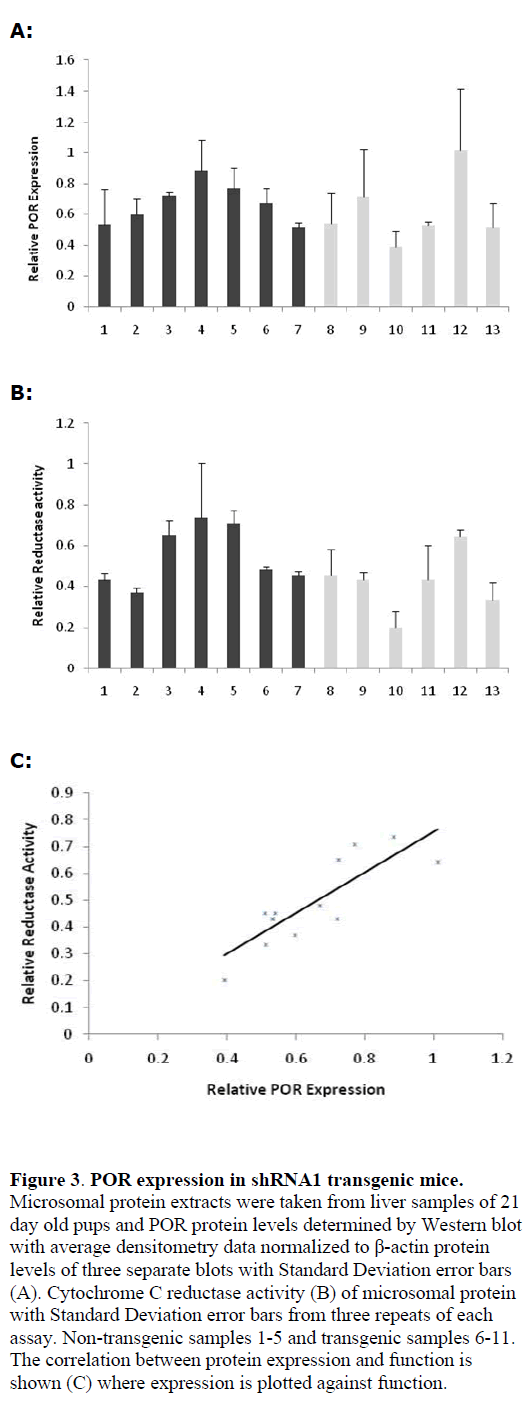 |
| Limited success has been noted when conjugated antisense oligonucleotides were delivered to the rat liver where a small reduction in POR reductase activity was seen (Venkateswaran et al, 2010). In this study the antisense oligos were delivered using a non-viral delivery system which allowed large doses of the oligonucleotides to be delivered into adult rats, and despite a modest reduction in function, no significant change was seen in the protein expression level. This, together with our findings, suggests that POR is hard to specifically knockdown in vivo using RNAi and antisense technologies, indicating that more factors may be involved than the just previously reported embryonic lethality. |
CONCLUSIONS |
| The overall aim of this study was to generate robust and versatile cell and animal systems that exhibited varying levels of POR activity, more accurately modelling the human population for pharmacological and toxicology studies. Specifically, altering POR activity would allow the role of cytochrome P450 activity on drug metabolism to be determined, enabling identification of bioavailability, efficacy and characterisation of drug metabolites to efficacy and toxicity to be evaluated (Henderson et al, 2003; Finn et al, 2007). |
| Although this study clearly demonstrates that both POR protein level and reductase activity can be significantly knocked down in Hepa-1 cells in vitro, the data also highlight the difficulty in knockdown of POR in an animal model. We believe that for strategies aiming to deliver gene knockdown in transgenic animals, the prior testing of RNAi constructs in cell systems is not necessarily a reliable indicator of subsequent in vivo activity. This study demonstrates that POR is a difficult target to knock down in vivo and inability to produce multi-copy transgenic animals suggests that a high level of shRNA expression is detrimental to the development of the embryo. |
ACKNOWLEDGMENTS |
| We thank Colin Henderson, John Hayes and Douglas Vasey for insightful discussion; and Colin Henderson for reagents. This work was funded through a Biotechnology and Biological Sciences Research Council (BBSRC)- CASE studentship to NM and the European Commission NEST029025 project INTEGRA. |
COMPETING INTERESTS |
| None declared. |
References |
|